Кора представляет собой слой серого вещества толщиной в среднем 3 мм. В кору приходят сенсорные волокна после «переключения» в таламусе, и из нее выходят моторные волокна, направляющиеся в спинной мозг.
Два мозговых полушария соединены между собой комиссурами - поперечными пучками нервных волокон. Главной из этих комиссур является толстая пластина мозолистого тела; она простирается спереди назад на 8 см и состоит более чем из 200 млн. нервных волокон, идущих из одного полушария в другое.
Кора каждого полушария образует шесть обособленных долей, разграниченных бороздами, из которых две особенно крупные - роландова и сильвиева. В передней части мозга выделяют лобную долю, в верхней - теменную, в боковой - височную, в задней - затылочную; под височной долей, в глубине сильвиевой борозды находится долька, называемая островком, а под мозолистым телом, на внутренней поверхности полушария-доля мозолистого тела (рис. А.24).
Рис. А.24. Кора большого мозга.
Между бороздами коры образуются валики, называемые извилинами, которые в большей или меньшей степени соответствуют областям с определенными функциями. Это могут быть сенсорные, моторные или ассоциативные зоны коры (см. рис. А. 19). Сенсорные зоны получают информацию от различных рецепторов, а моторные зоны посылают команды, управляющие движениями. Таким образом, сенсорные области мозговой коры представляют собой конечные пункты на пути волокон, связанных с периферической нервной системой, и их разрушение приводит к потере чувствительности в той области тела, где расположены соответствующие рецепторы. Моторные области дают начало волокнам, разрушение которых вызывает паралич конечности, управляемой нейронами соответствующей области коры.
Наиболее значительную часть коры, однако, занимают ассоциативные зоны, организация которых наиболее характерна для этой мозговой структуры. По сути дела, именно эти зоны, лишенные какой-либо явной специализации, ответственны за объединение и переработку информации и программирование действий. Благодаря этому они составляют основу таких высших процессов, как память, научение, мышление и речь (см. документ 8.4).
А. Сенсорные зоны. Такие зоны имеются в разных долях коры. Зона общей чувствительности находится в теменной доле, зрительная зона - в затылочной, слуховая - в височной, вкусовая - в нижней части теменной доли, а обонятельная в двух обонятельных луковицах, находящихся под большим мозгом.
Зона общей чувствительности расположена в извилине, идущей вдоль роландовой борозды, в теменной доле и получает сигналы от рецепторов кожи. Все тело человека - головой вниз, а пальцами ног вверх - представлено здесь в виде областей (проекций), поверхность которых пропорциональна чувствительности соответствующих частей тела; так, проекция кисти намного больше проекций спины или ног (рис. А.25).

Рис. А.25. Величина проекций сенсорных волокон в соместетической зоне коры несоразмерна с величиной тех участков тела, от которых эти волокна отходят (А). То же самое относится и к распределению центров моторной зоны, ведающих произвольными движениями (Б). Изобразив проекции различных частей тела в коре, эту несоразмерность можно иллюстрировать в виде сенсорного или моторного гомункулюса.
Повреждение всей этой зоны или какой-либо ее части приводит к блокаде сенсорных сигналов от соответствующих областей тела; в результате здесь исчезают тактильные, температурные и болевые ощущения, хотя внешние стимулы продолжают возбуждать рецепторы кожи и вызывать поток импульсов в идущих от них нервных путях.
Ассоциативная зона, находящаяся в верхней части теменной области, является гностической и отвечает за узнавание и восприятие стимулов, вызвавших ощущения на уровне теменной извилины.
Зона зрительной чувствительности расположена в затылочной доле вдоль шпорной борозды, и информация, передаваемая каждой ганглиозной клеткой сетчатки, очень точно проецируется в разные ее точки.
Затылочная зона каждого полушария мозга получает информацию от противоположной половины поля зрения. Прежде чем войти в большой мозг, часть волокон обоих зрительных нервов перекрещивается, образуя так называемую зрительную хиазму (рис. А.26). В результате этого перекрещивания левая зрительная доля получает волокна от обоих глаз, несущие информацию о правой половине поля зрения, а правая доля - о левой половине. Таким образом, в результате интеграции нервных сигналов от обеих сетчаток в мозгу воссоздается трехмерный образ предмета, изображения которого на правой и левой сетчатках несколько различны.

Рис. А.26. Зрительный перекрест (хиазма) и зрительные пути. Информация о событиях в правой половине поля зрения поступает в левую затылочную долю из левой части каждой сетчатки; информация же о правой половине поля зрения направляется в левую затылочную долю из правых частей обеих сетчаток. Такое перераспределение информации от каждого глаза происходит в результате перекрещивания части волокон зрительного нерва на уровне зрительной хиазмы.
Зрительное восприятие предметов, слов и чисел осуществляется в ассоциативной зоне, расположенной вокруг сенсорной зоны.
Зона слуховой чувствительности находится в височной области коры. Каждая из двух височных долей получает информацию, улавливаемую обоими ушами. Поэтому даже значительное повреждение слуховой зоны не может привести к глухоте, если оно, конечно, не затрагивает обоих мозговых полушарий.
Восприятие звуков, включая интерпретацию слов и мелодий, происходит в ассоциативной зоне, находящейся под сенсорной зоной (см. документ 8.4).
Вкусовая и обонятельная чувствительность локализована в зонах, расположенных сравнительно недалеко друг от друга. Зона вкусовой чувствительности находится в основании восходящей извилины и отвечает за расшифровку нервных сигналов, приходящих от языка. Доминирующая у большинства животных зона обонятельной чувствительности редуцирована у человека до двух обонятельных луковиц, являющихся продолжением обонятельных полосок в основании большого мозга.
Б. Двигательные (моторные) зоны. Область, ведающая произвольными движениями, расположена в извилине лобной доли, тянущейся вдоль роландовой борозды. Выходящие из нее моторные волокна направляются в спинной мозг либо прямо, проходя в виде двух пучков через варолиев мост и продолговатый мозг (где перекрещиваются), либо непрямым путем - через мозжечок и различные ядра, ответственные за координацию движений.
Как и в зоне общей чувствительности, в моторной зоне в виде проекций представлено все тело человека (головой вниз, пальцами ног вверх); площадь этих проекций пропорциональна сложности управления соответствующими группами мышц (см. рис. А.25, Б ).
Ассоциативная зона, прилегающая к моторной области и тесно взаимодействующая с расположенным под ней полосатым телом (см. выше), ответственна за моторные автоматизмы, а также за программирование и координацию более сложных и тонких движений. Повреждения этой зоны сопровождаются расстройством, получившим название двигательной апраксии (см. документ 8.4).
В. Зоны мышления и планирования действий. Собственно говоря, зон, где «рождаются» мысли, не существует. В принятии даже самого незначительного решения участвует весь мозг. В действие вступают разнообразные процессы, происходящие как в различных зонах коры, так и в низших нервных центрах.
Многообразны и формы самого процесса мышления. Он может быть направлен на решение самых различных задач - от простой оценки пространственных или временных отношений до предвидения результатов действий - и, помимо прочего, может быть связан с функциями памяти и речи или даже с владением сложными психомоторными навыками (см. дополнение А.3).
В любой момент времени наш мозг осведомлен о положении тела в пространстве благодаря той информации, которая поступает в него по различным сенсорным каналам. Эта информация, по-видимому, стекается в область, расположенную на стыке трех долей мозга, включающих главные сенсорные зоны. Речь идет о так называемой «дугообразной складке», расположенной в верхней части сильвиевой борозды (см. рис. А.24), которая получает также нервные сигналы, передаваемые таламусом и различными ядрами. Повреждение этой зоны ведет к расстройству жестикуляции и ориентировки в пространстве.
Способность мозга определять время совершения события в основном зависит от памяти. Проведенные недавно исследования, по всей видимости, указывают на то, что способность ориентироваться во времени особенно свойственна высшим животным и что она в известных пределах не зависит от циркадианных ритмов (Richelle, Lejeune, 1986).
Память, очевидно, не связана с какой-то одной специфической областью мозга; она зависит от многочисленных зон, играющих важную роль. В особенности это касается некоторых областей височной коры и в еще большей"мере - гиппокампа (см. документ 8.1).
Речь и язык одновременно связаны с такими сенсорными функциями, как слух и зрение, и с двигательными функциями, необходимыми для устной речи и письма (см. документ 8.4). Центры, ответственные за эти функции, находятся в разных областях мозга, особенно в лобной, затылочной и височной долях. У подавляющего большинства людей лингвистическая активность контролируется левым полушарием мозга.
Планирование действий, которое, собственно, и составляет суть мышления, происходит в префронтальной коре (т. е. в передних участках лобных долей) в результате объединения и переработки ею информации, получаемой и расшифровываемой в других зонах коры. Именно в префронтальной коре находятся структуры, определяющие способность к счету, предсказанию и предвидению*.
* У человека эта область занимает 29% поверхности коры, у шимпанзе 17%, а у собаки всего 7% (Changeux).
Наконец, управление сложными психомоторными функциями осуществляется на уровне верхних отделов мозгового ствола. Эта область мозга представляет собой настоящую «телефонную станцию» (Lazorthes, 1973), объединяющую информацию от рецепторов и моторные сигналы из коры мозга. Благодаря этому она может контролировать выполнение движений, планируемых лобной корой.
Специализация полушарий
Развитие центральной нервной системы уже у плоских червей (например, у планарий) сопровождается возникновением билатеральной (двусторонней) симметрии всего тела. Тело оказывается разделенным в продольном направлении на две половины, каждая из которых представляет собой зеркальное отражение другой, причем левая половина тела находится под контролем правой стороны мозга, и наоборот.
В процессе эволюции предков человека каждое мозговое полушарие приобретало все большую специализацию, что в особенности проявилось в предпочтительном пользовании правой или левой рукой, развитии речи, пространственной ориентации и полярности эмоциональных состояний.
Предпочтительное пользование той или другой рукой. Правши составляют около 90% всех людей; по-видимому, доминирование правой руки существовало уже у пещерных предков человека*. Не следует, однако, думать, что такая ситуация обязательно обусловлена наследственными факторами. Статистически установлено, что ребенок, у которого оба родителя левши, имеет примерно один шанс из двух стать правшой.
* По-видимому, при создании наскальных изображений человека контур руки нередко наносился с помощью трафарета, которым служила свободная рука самого художника, и в 80% таких случаев это была левая рука. Значит, контур обводился обычно правой рукой.
Речь. У подавляющего большинства людей центры речи расположены в левом полушарии. Только 5% правшей и 30% левшей, т.е. менее 8% всех людей, разговаривают с помощью правого полушария. Согласно Рош-Лекуру (цит. по Changeux, 1983), все дети появляются на свет с речевыми зонами в обоих полушариях, однако в процессе развития на первом году жизни одно из них «берет верх» над другим. Поэтому отсутствие или случайная утрата одного полушария при рождении или в первые два года жизни может быть компенсирована, так как соответствующие функции способно взять на себя второе полушарие.
То, что некоторые функции представлены только в одном полушарии, может означать, что это полушарие (обычно левое) подавляет активность другого. Иными словами, вследствие блокады недоминантного полушария доминантным через межполушарные волокна мозолистого тела недоминантное полушарие остается пассивным.
В дополнении А.З приводятся наблюдения ученых за работой обоих мозговых полушарий, ставших независимыми после перерезки мозолистого тела. Эти наблюдения позволили выявить важную роль мозолистого тела в межполушарных взаимодействиях и в особенности роль доминантного полушария в объединении информации. Благодаря такой организации большого мозга вся нервная система в целом получает возможность работать согласованно и эффективно. Так, например, нервные сигналы, вызванные раздражением левой руки и приходящие в правое полушарие, автоматически передаются в доминантное левое полушарие. Лишь после того как левое полушарие ознакомилось с этой информацией, в первое полушарие посылается команда, заставляющая левую руку выполнить нужное движение.
Эмоциональные состояния. По-видимому, каждое полушарие мозга, помимо прочего, отвечает за направленность чувств человека и их позитивную или негативную окраску. Так, например, если патологический очаг у больного эпилепсией находится в левом полушарии мозга, человека нередко охватывает беспредметный смех, а если в правом, то больной более склонен к грусти и слезам.
Было также показано, что у людей во время депрессии в области правого полушария нередко регистрируются аномальные электрические волны. Это привело к предположению, что правое полушарие ответственно за эмоциональные состояния с негативной окраской и способствует тому, что человек видит прежде всего отрицательные стороны событий, тогда как левое полушарие придает эмоциональным реакциям на те или иные события положительную окраску. Таким образом, чувство или эмоциональное состояние человека будет определяться балансом этих противоположных тенденций. Однако, как подчеркивает Шанжё, вопрос о том, как мозгу удается без острого конфликта сделать взвешенный выбор, до сих пор остается полной загадкой.
Половые различия. Были обнаружены некоторые различия в строении мозга у мужчин и женщин. Например, недавно выяснилось, что у женщин в определенном участке мозолистого тела больше нервных волокон, чем у мужчин. Это может означать, что межполушарные связи у женщин более многочисленны и поэтому у них лучше происходит объединение информации, имеющейся в обоих полушариях; этим можно объяснить и некоторые половые различия в поведении. Кроме того, выявленные у женщин более высокие показатели, связанные с лингвистическими функциями, памятью, аналитическими способностями и тонким ручным манипулированием можно связать с большей относительной активностью у них левого полушария мозга. Напротив, функции восприятия и способность к оценке пространственных отношений и художественному творчеству, видимо, лучше развиты у мужчин, что может объясняться большим участием в этих процессах правого полушария. Еще раз, однако, отметим, что в первые годы жизни оба полушария способны хранить одинаковые количества и одинаковые виды информации и что специализация полушарий происходит лишь очень постепенно. В связи с этим можно задаться вопросом: какова роль культуры и воспитания в формировании различий между женщинами и мужчинами, в частности различий в развитии нервных функций, обусловливающих те или иные способности?
Слуховая сенсорная система
Функция слуховой системы состоит в формировании слуховых ощущений человека в ответ на действие звуковых волн, представляющих собой распространяющиеся колебания молекул воздуха (упругой среды). Периферическая часть слуховой системы включает наружное, среднее и внутреннее ухо, в котором расположены слуховые рецепторы. Ее центральную часть образуют проводящие пути, переключательные ядра и слуховая кора, расположенная в обоих полушариях в глубине латеральной борозды, отделяющей височную долю от лобной и передних отделов теменной доли.
Кортиев орган
Кортиев орган – рецепторная часть слухового анализатора, расположенная внутри перепончатого лабиринта. В процессе эволюции возникает на основе структур органов боковой линии. Воспринимает колебания волокон, расположенных в канале внутреннего уха, и передаёт в слуховую зону коры больших полушарий, где и формируются звуковые сигналы. В кортиевом органе начинается первичное формирование анализа звуковых сигналов. Кортиев орган располагается в спирально завитом костном канале внутреннего уха – улитковом ходе, заполненном эндолимфой и перилимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке.
Волосковые клетки
Внутренние волосковые клетки располагаются в один ряд; общее число их по всей длине перепончатого канала достигает 3500. Наружные волосковые клетки располагаются в 3–4 ряда; общее число их составляет 12 000–20 000. Каждая волосковая клетка имеет удлиненную форму. Один полюс клетки фиксирован на основной мембране; второй ее полюс находится в полости перепончатого канала улитки. На конце этого полюса рецепторной клетки имеются волоски: их число на каждой внутренней клетке составляет 30–40 и они очень короткие – 4–5 мкм (4–5-10-6 м); на каждой наружной клетке число волосков достигает 65–120, они тоньше и длиннее. Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной пластинкой, или текториальной мембраной, которая по всему ходу перепончатого канала расположена над волосковыми клетками. При действии звуков основная мембрана начинает колебаться, волоски рецепторных клеток касаются текториальной мембраны и деформируются. Это вызывает генерацию электрических потенциалов, а затем через синапсы – возбуждение волокон слухового нерва. Электрические явления в улитке. При отведении электрических потенциалов от разных частей улитки исследователи обнаружили 5 различных электрических феноменов. Два из них – мембранный потенциал слуховой рецепторной клетки и потенциал эндолимфы – не обусловлены действием звука (они наблюдаются и при отсутствии звуковых раздражений). Три электрических явления – микрофонный потенциал улитки, суммационный потенциал и потенциалы слухового нерва – возникают под влиянием звуковых раздражений
Механизм возбуждения кортиева органа
Физиологический механизм восприятия звука основан на двух процессах, происходящих в улитке:
1) разделение звуков различной частоты по месту их наибольшего воздействия на основную мембрану улитки
2) преобразование рецепторными клетками механических колебаний в нервное возбуждение.
Звуковые колебания, поступающие во внутреннее ухо через овальное окно, передаются перилимфе, а колебания этой жидкости приводят к смещениям основной мембраны. От высоты звука зависит высота столба колеблющейся жидкости и, соответственно, место наибольшего смещения основной мембраны. Таким образом, при различных по высоте звуках возбуждаются разные волосковые клетки и разные нервные волокна. Увеличение силы звука приводит к увеличению числа возбужденных волосковых клеток и нервных волокон, что позволяет различать интенсивность звуковых колебаний. Преобразование колебаний в процесс возбуждения осуществляется специальными рецепторами – волосковыми клетками. Волоски этих клеток погружены в покровную мембрану. Механические колебания при действии звука приводят к смещению покровной мембраны относительно рецепторных клеток и изгибанию волосков. В рецепторных клетках механическое смещение волосков вызывает процесс возбуждений.
Теория бегущей волны (или теория Бекеша)
Согласно гидродинамической теории Бекеши и Флетчера, под влиянием звуков в лимфе улитки происходят сложные динамические процессы и деформации мембран. При быстрых колебаниях подножной пластинки относительно большая инерция столба лимфы в обеих лестницах не позволяет ему следовать за быстрыми колебаниями стремени.
Это ведет к повышению давления в вестибулярной лестнице, вследствие чего вестибулярная мембрана, а затем и базилярная пластинка прогибаются, при э том давление передается дальше на лимфу барабанной лестницы и на мембрану окна улитки.
Чем выше звук, тем ближе к окну улитки прогиб базилярной пластинки. Самые низкие звуки вызывают деформацию ее у верхушки улитки. В настоящее время основной считается теория пространственного расположения звуков. По новейшим экспериментальным данным Бекеши и Флетчера, место максимальной деформации соответствует пространственному расположению звуков на основной мембране, где наблюдались вихревые движения лимфы.
Слуховой нерв
Слуховой нерв – восьмой черепно-мозговой чувствительный нерв (VIII), по которому от внутреннего уха к головному мозгу поступают импульсы от вестибулярного аппарата и кортиева органа внутреннего уха. Этот нерв имеет две части: По нерву улитки передаются импульсы от слухового (нерв преддверия и нерв улитки) органа улитки; это слуховой нерв. Нерв преддверия, по которому проходят импульсы от полукружных каналов, маточек и мешочков, передавая в головной мозг информацию о положении тела и осуществляемых им движениях, это нерв равновесия.
Роль подкорковых ядер в восприятии слуха
К подкорковым ядрам относятся хвостатое ядро, бледный шар и скорлупа. Они находятся в толще больших полушарий головного мозга, между лобными долями и промежуточным мозгом. Эмбриональное происхождение хвостатого ядра и скорлупы едино, поэтому о них говорят иногда как о едином – полосатом теле (стриатум). Бледный шар, филогенетически наиболее древнее образование, обособлен от полосатого тела и морфологически, и функционально.
Функции базальных ганглиев по сравнению с функциями других образований нервной системы менее изучены. Это обусловлено труднодоступностью их для прямого исследования, при сохранении целостности мозга. Разрушение отдельных частей полосатого тела приводит к нарушению обширных нервных связей подкорковых ядер с корой и ядрами стволовой части мозга.
К настоящему времени получено достаточно экспериментальных фактов, позволяющих говорить, что логичная, хорошо построенная система о стриатуме и бледном шаре как высших подкорковых двигательных центрах – хранителях древних автоматизмов не отвечает действительности. Следовательно, и сложившиеся ранее представления о физиологических механизмах расстройств, вызываемых нарушением функций стриатума или бледного шара, являются неверными.
Слуховая кора
Существует две отдельные части слуховой коры: первичная слуховая кора и ассоциативная слуховая кора (называемая также вторичной слуховой корой). Первичная слуховая кора непосредственно возбуждается проекциями от медиального коленчатого тела, тогда как ассоциативная слуховая кора возбуждается вторично импульсами из первичной слуховой коры, а также проекциями из таламических ассоциативных зон, прилежащих к медиальному коленчатому телу.
Восприятие частоты звука в первичной слуховой коре. В первичной слуховой коре и ассоциативной слуховой коре обнаружены, по крайней мере, шесть тонотопических карт. В каждой из этих карт высокочастотные звуки возбуждают нейроны у одного конца карты, а низкочастотные звуки – у противоположного конца. В основном, низкочастотные звуки локализуются спереди, а высокочастотные – сзади.
Это справедливо не для всех карт. Возникает вопрос: почему слуховая кора имеет так много разных карт? Ответ, вероятно, в том, что каждая из отдельных областей анализирует одно из специфических свойств звука. Например, одна из больших карт в первичной слуховой коре, вероятно, различает сами звуковые частоты и дает человеку физическое ощущение высоты звуков. Другая карта, вероятно, используется для определения направления, откуда исходит звук.
Другие области слуховой коры выделяют особые качества, например внезапное начало звуков, или, возможно, особые модуляции, например выделение звуков определенной частоты из шума.
Диапазон частот, на которые реагирует каждый нейрон слуховой коры, гораздо уже, чем в улитке и релейных ядрах мозгового ствола. Вновь обратившись к рисунку, можно видеть, что базальная мембрана улитки стимулируется звуками всех частот, и такое же широкое звуковое представительство обнаруживается в улитковых ядрах. Однако когда возбуждение достигает мозговой коры, большинство звукочувствительных нейронов реагируют лишь на узкий, а не на широкий диапазон частот.
Следовательно, где-то по пути механизмы анализа «обостряют» реакцию на частоту. Полагают, что причиной этого обостряющего эффекта является главным образом феномен латерального торможения. Это значит, что стимуляция улитки одной частотой тормозит звуковые частоты с обеих сторон этой первичной частоты; причиной являются коллатеральные волокна, ответвляющиеся от первичного сигнального пути и оказывающие тормозное влияние на прилежащие пути. Важность такого эффекта показана также для усиления особенностей соматосенсорных, зрительных и других типов ощущений.
Многие нейроны слуховой коры, особенно в ассоциативной слуховой коре, не просто реагируют на специфические звуковые частоты в ухе. Полагают, что эти нейроны «связывают» разные звуковые частоты друг с другом или звуковую информацию с информацией от других сенсорных областей коры. Действительно, теменная часть ассоциативной слуховой коры частично перекрывает соматосенсорную область II, что, вероятно, обеспечивает возможность ассоциации слуховой информации с соматосенсорной.
Особенности слуха человека
Человеческое ухо способно воспринять звуки с частотой от 20 до 22000 Гц, но его чувствительность не является одинаковой в этом интервале. Она зависит от частоты звука. Эксперименты указывают на то, что в тихой окружающей обстановке чувствительность уха максимальна при частотах от 2 до 4 кГц. Стоит отметить, что частотный диапазон человеческого голоса также весьма ограничен. Он располагается в интервале от 500 Гц до 2 кГц. Существование порога слышимости дает основу для построения методов сжатия звука с потерями. Можно удалять все сэмплы, величина которых лежит ниже этого порога. Поскольку порог слышимости зависит от частоты, кодер должен знать спектр сжимаемого звука в каждый момент времени. Для этого нужно хранить несколько предыдущих входных сэмплов (обозначим это число п – 1; оно или фиксировано, или задается пользователем). При вводе следующего сэмпла необходимо на первом шаге сделать преобразование п сэмплов в частотную область. Результатом служит вектор, состоящий из га числовых компонент, которые называются сигналами. Он определяет частотное разложение сигнала. Если сигнал для частоты / меньше порога слышимости этой частоты, то его следует отбросить.
Для эффективного сжатия звука применяются еще два свойства органов слуха человека. Эти свойства называются частотное маскирование и временное маскирование. Частотное маскирование (его еще называют слуховое маскирование) происходит тогда, когда нормально слышимый звук накрывается другим громким звуком с близкой частотой. Этот звук приподнимает порог слышимости в своей окрестности (пунктирная линия). В результате звук, обозначенный тоненькой стрелкой в точке «х» и имеющий нормальную громкость выше своего порога чувствительности, становится неслышимым; он маскируется более громким звуком. Хороший метод сжатия звука должен использовать это свойство слуха и удалять сигналы, соответствующие звуку «х», поскольку они все равно не будут услышаны человеком. Это один возможный путь сжатия с потерями. Частотное маскирование (область под пунктирной линией на рис.) зависит от частоты сигнала. Оно варьируется от 100 Гц для низких слышимых частот до более чем 4 кГц высоких частот. Следовательно область слышимых частот можно разделить на несколько критических полос, которые обозначают падение чувствительности уха (не путать со снижением мощности разрешения) для более высоких частот.
Можно считать критические полосы еще одной характеристикой звука, подобной его частоте. Однако, в отличие от частоты, которая абсолютна и не зависит от органов слуха, критические полосы определяются в соответствии со слуховым восприятием. В итоге они образуют некоторые меры восприятия частот. Критические полосы можно описать следующим образом: из-за ограниченности слухового восприятия звуковых частот порог слышимости частоты приподнимается соседним звуком, если звук находится в критической полосе. Это свойство открывает путь для разработки практического алгоритма сжатия аудиоданных с потерями. Звук необходимо преобразовать в частотную область, а получившиеся величины (частотный спектр) следует разделить на под полосы, которые максимально приближают критические полосы. Если это сделано, то сигналы каждой из подполос нужно квантовать так, что шум квантования (разность между исходным звуковым сэмплом и его квантованными значениями) был неслышимым.
Еще один возможный взгляд на концепцию критической полосы состоит в том, что органы слуха человека можно представить себе как своего рода фильтр, который пропускает только частоты из некоторой области (полосы пропускания) от 20 до 20000 Гц. В качестве модели ухо-мозг мы рассматриваем некоторое семейство фильтров, каждый из которых имеет свою полосу пропускания. Эти полосы называются критическими. Они пересекаются и имеют разную ширину. Они достаточно узки (около 100 Гц) в низкочастотной области и расширяются (до 4-5 кГц) в области высоких частот. Ширина критической полосы называется ее размером. Для измерения этой величины вводится новая единица «барк» («Вагк» от H.G. Barkhausen). Один барк равен ширине (в герцах) одной критической полосы. Временное маскирование происходит, когда громкому звуку А частоты / по времени предшествует или за ним следует более слабый звук В близкой частоты. Если интервал времени между этими звуками не велик, то звук В будет не слышен. Рис. 6.6 иллюстрирует пример временного маскирования. Порог временного маскирования от громкого звука в момент времени 0 идет вверх сначала круто, а потом полого. Более слабый звук в 30 дБ не будет слышен, если он раздастся за 10 мл/сек до или после громкого звука, но будет различим, если временной интервал между ними будет больше 20 м/сек.
Кора головного мозга
Кора больших полушарий представляет собой слой нейронов толщиной около 3 мм. Кора каждого полушария образует шесть обособленных долей, разграниченных бороздами. Лобная, теменная, височная и затылочная доли хорошо видны на рис. 3.11, а островок и доля мозолистого тела спрятаны на внутренних поверхностях больших полушарий. В коре больших полушарий выделяют зоны трех типов.
Рис. 3.11.
1. Сенсорные зоны, которые получают информацию от рецепторов. Они имеются в разных долях коры. Зона общей чувствительности находится в теменной доле, зрительная зона – в затылочной, слуховая – в височной, вкусовая – в нижней части теменной доли. Повреждение какой-либо из этих зон приводит к блокаде сенсорных сигналов от соответствующих органов чувств. При поступлении сенсорной информации поток нервных импульсов достигает коры. Например, зрительные проекционные зоны активизируются при анализе зрительной информации (рис. 3.12).

Рис. 3.12.
2. Моторные зоны, которые посылают в нижележащие структуры команды, управляющие движениями. Область, отвечающая за произвольные движения, расположена в лобной доле. Отходящие от нее волокна направляются в спинной мозг и оттуда – к мышцам.
Сенсорные и моторные зоны называют проекционными, так как они формируют своеобразные карты, на которых специфические участки коры соответствуют частям тела, которыми они управляют или информацию от которых получают (см. рис. в гл. 7).
3. Ассоциативные зоны, занимающие до 75% площади коры головного мозга человека. Данные области коры больших полушарий можно охарактеризовать как "неспецифические". Эти участки коры не задействованы в выполнении первичных моторных и сенсорных функций. Долгое время считалось, что ассоциативные зоны коры – это балласт мозга. Отсюда, кстати, возникло расхожее и крайне живучее мнение о том, что в повседневной жизни человек использует лишь малую часть потенциала мозга. Па самом деле в этих зонах идет наиболее кропотливая обработка информации. Ассоциативные зоны представляют особый интерес для психологов, так как они принимают участие в протекании высших "корковых" процессов: восприятия, памяти, мышления, воображения, речи и т.д. Например, ассоциативные зоны лобных долей необходимы при планировании действий, повреждение этих зон мозга делает человека неспособным к выходу за пределы конкретной ситуации, он начинает жить исключительно "здесь и сейчас", хотя в остальном его умственные способности остаются неизменными. Установлено, что при поражении зоны Брока (она находится в передней лобной доле левого полушария) человек теряет способность правильно произносить слова, а при поражении зоны Вернике (находится в височной доле левого полушария) – понимать обращенную к нему речь.
Важно отмстить, что в мозге нет конкретной структурной организации, которая соответствовала бы категориям нашего поведения или субъективным состояниям. Так, в ассоциативной коре нет участков, где рождаются, например, мысли или воспоминания. Даже в осуществление простых психических функций вовлечено множество структур мозга. В случае же сложных психических процессов уместнее говорить о вариативных функциональных системах, обслуживающих то или иное действие.
Проблема локализации психики.
Теория системной динамической локализации высших психических функций А. Р. Лурии
В подходах к тому, как определить "место" психических функций в мозге, существует две крайних позиции: локализационизм и антилокализационизм. Представители первого подхода считают, что за осуществление каждой психической функции отвечает строго определенный участок мозга. Соответственно, при локальных поражениях мозга психические функции будут страдать избирательно. Ярким воплощением локализационизма является френологическая концепция Ф. Галля, согласно которой каждый "бугорок" мозга отвечал за конкретную умственную способность (включая и такие комплексные способности, как "любовь к родителям" или "честолюбие"), а по их выраженности можно было судить о психологических особенностях человека. Представители второго подхода уверены, что психические функции являются результирующей работой всего мозга в целом. Согласно этой точке зрения при поражении мозга страдает вся психика, и чем больше площадь поражения, тем серьезнее нарушение.
Истина лежит посередине. Когда речь идет о примитивных сенсорных и моторных процессах, которые жестко генетически связаны с проекционными зонами коры больших полушарий, наиболее адекватным представляется первый подход. Такие элементарные процессы, как светоразличительная способность сетчатки глаза, различение звуков или тактильные ощущения, за миллионы лет эволюции обрели свое место на определенной ограниченной группе нервных клеток. История человеческой культуры насчитывает всего десятки тысяч лет, так что за это время просто не могли сформироваться особые, генетически закрепленные "зоны счета", "зоны чтения", "зоны письма", "зоны музыки" и т.п.
По концепции А. Р. Лурии, высшие психические функции представляют собой функциональные системы, каждая из которых включает в себя несколько звеньев. В данном контексте трактовка термина "функция" отличается от обыденной: функция как действие какого-либо органа (например, функция слезной железы – выделение слезной жидкости). Функция понимается здесь как сложная деятельность, направленная на решение задачи. В то время как конечная задача функциональной системы является постоянной (например, "узнать, который час"), ее составные части пластичны (можно посмотреть на часы, или позвонить по телефону, или спросить у прохожего).
Сам термин "функциональная система " был введен П. К. Анохиным в 1935 г. для описания организации процессов в целостном организме, взаимодействующем со средой. Поясним это понятие па примере достаточно простого процесса – дыхания. Известно, что определенная область продолговатого мозга раздражима по отношению к концентрации углекислого газа в крови. Если его уровень повышается, посылается импульс в спинной мозг и оттуда – к мышцам диафрагмы, которые в ответ сокращаются (вдох). Однако если перерезать двигательный нерв диафрагмы, животное не задохнется. Вдох будет осуществлен за счет межреберных мышц. Исключение межреберных мышц приведет к использованию других способов доставки воздуха в легкие, например заглатыванию воздуха. Иначе говоря, звенья, входящие в данную функциональную систему, не представляют собой фиксированной и постоянной цепи реакций и обладают значительной замещаемостью. Для объяснения работы психических функций в указанном смысле А. Р. Лурия предложил теорию динамической системной локализации высших психических функций: "Если дыхание представляет собой столь сложную и пластичную функциональную систему, совершенно естественно, что нс может быть и речи о локализации этой функции в каком-либо ограниченном участке мозга".
По мысли ученого, структуры мозга, в первую очередь ассоциативные зоны коры, "функционально многозначны", т.е. могут включаться в различные функциональные системы. Высшие психические функции социальны по своему происхождению и формируются прижизненно. В процессе их формирования складываются "функциональные мозговые органы" (А. Н. Леонтьев), объединяющие в единые системы множество специфичных участков мозга. Причем на последовательных этапах своего развития высшие психические функции не сохраняют единой структуры, но решают одни и те же задачи с помощью различных закономерно сменяющих друг друга систем связей. Например, мозговая организация речи у ребенка, не владеющего грамотой, значительно отличается от таковой у грамотного взрослого. Участие слуховых и зрительных зон коры, существенное на ранних этапах формирования познавательной деятельности, перестает играть такую роль на поздних этапах, когда мышление начинает опираться на согласованную деятельность разных систем коры мозга. Поэтому поражение, например, зрительных сенсорных отделов коры в раннем детстве приводит к недоразвитию познавательных способностей и мышления, в то время как у взрослого аналогичное поражение может компенсироваться влиянием уже сформировавшихся высших функциональных систем. Более того, различия в специализации различных областей мозга в зависимости от особенностей социальных воздействий можно обнаружить даже на таких фундаментальных явлениях, как явления межполушарной асимметрии (см. ниже). Если у европейцев, чья письменность носит фонематический характер (мы пишем, условно говоря, "звуки"), в процессы, связанные с речью, вовлечено преимущественно левое полушарие мозга, то у народов, обладающих символьной письменностью, например у китайцев, – и левое, и правое практически в равной мере.
Огромный успех концепции динамической системной локализации высших психических функций А. Р. Лурии связан в первую очередь с возможностями ее эффективного использования в клинической практике. Если какое-то звено функциональной системы поражено, представляется возможность перестроить систему и добиться компенсации функции.
В качестве иллюстрации рассмотрим страшное заболевание, возникающее обычно в результате кровоизлияния в мозг, – афферентную моторную афазию. Симптоматика этого недуга заключается в том, что больной не в состоянии правильно произносить слова, путая близкие по артикуляции звуки, например "л" и "н", "б" и "м" и т.д. Случается это, если поражена зона Брока, расположенная в левой лобной доле коры больших полушарий мозга. Программа восстановления речевой функции в данном случае направлена на создание новых систем движений для произнесения звуков. То, что обычный человек делает автоматически, здесь приходится выстраивать как отдельный навык (вспомните модель Н. А. Бернштейна). При наблюдении за такими больными выясняется, что вне речевой ситуации они способны совершать необходимые движения. Например, артикуляторная картина звуков "п" и "б" сходна с той, которая возникает при резком задувании спички. Положение губ в момент, когда человек дует на горячий чай, такое же, как и при произнесении звука "у", и т.д. Больной много раз дует па спичку и старается осознать и зафиксировать те изменения, которые происходят. С помощью различных внешних средств (зеркал, особых схем) добиваются закрепления заново выученных звуков. Таким образом, в результате кропотливой реабилитационной работы высшие сознательные психические функции замещают низшие автоматизмы.
«Мозг – это последняя из тайн природы,
которая когда-либо откроется человеку.»
англ. физиолог Чарлз Скотт Шеррингтон.
«Асимметрия является основным
свойством жизни.»
Луи Пастер.
Большие полушария – парные образования головного мозга. У человека они достигают ≈ 80% от общей массы мозга. Большие полушария осуществляют регуляцию высших нервных функций, лежащих в основе всех психических процессов человека, в то время как стволовая часть мозга обеспечивает низшие функции нервной системы, связанные с регуляцией деятельности внутренних органов.
Высшие функции обеспечиваются деятельностью особого отдела больших полушарий – коры головного мозга , которая несет главную ответственность за формирование условно-рефлекторных реакций. У человека по сравнению с животными кора одновременно ответственна и за согласование работы внутренних органов. Такое возрастание роли коры в регуляции всех функций в организме называется кортикализацией функций .
Кора выполняет следующие функции :
1 – взаимодействие организма с внешней средой за счет безусловных и условных рефлексов.
2 – осуществление высшей нервной деятельности (поведения) организма.
3 – выполнение высших психических функций (мышления и сознания).
4 – регуляция работы внутренних органов и обмена веществ в организме.
Кора больших полушарий представлена 12-18 миллиардами клеток, расположенных тонким слоем 3-4 мм на площади 2400 см 2 . 65-70 % этой площади находится в глубине борозд, а 30-35 % - на видимой поверхности полушарий. Кора состоит из нервных клеток, их отростков и нейроглинов, для которых характерно обилие межнейронных связей.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Все нейроны вертикальной колонки отвечают на одно и тоже афферентное раздражение одинаковой реакцией и совместно формируют эфферентный ответ. Распространение возбуждения в горизонтальном направлении (иррадиация) обеспечивается поперечными волокнами, идущими от одной вертикальной колонки к другой, а ограничивается – процессами торможения. Возникновение возбуждения в вертикальной колонке нейронов приводит к активности спинальные мотонейроны и к сокращению связанных с ними мышц.
Упорядоченное положение клеток в коре называется цитоархитектоникой , а их волокон – миелоархитектоникой .
При микроскопическом исследовании в коре различают шесть слоев нервных клеток:
1 – молекулярный (горизонтально расположенные клетки и волокна + дендриты пирамидных клеток),
2 – наружный зернистый (звездчатые и мелкие пирамидные клетки + тонкие нервные волокна),
3 – наружный пирамидный (средние и малые пирамидные клетки + восходящие волокна),
4 – внутренний зернистый (звездчатые клетки + таламо-корковые волокна и горизонтальные миелиновые волокна),
5 – внутренний пирамидный (крупные пирамидные клетки Беца от которых начинаются пирамидные проводящие пути),
6 – мультиформный (мелкие полиморфные клетки).
В первом слое коры волокна образуют полоску молекулярной пластинки . Во втором слое залегают тонкие волокна наружной зернистой пластинки . В составе четвертого слоя коры находится полоска внутренней зернистой пластинки (наружная полоска Байярже). В пятом слое содержатся волокна внутренней пирамидной пластинки (внутреняя полоска Байярже).
Основная информация в кору поступает по специфическим афферентным проводящим путям, заканчивающимся на клетках 3 и 4 слоев. Неспецифические пути от РФ заканчиваются в верхних слоях коры и регулируют ее функциональное состояние (возбуждение, торможение).
Звездчатые нейроны выполняют главным образом чувствительную (афферентную) функцию. Пирамидные и веретеновидные клетки – это преимущественно двигательные (эфферентные) нейроны.
Часть клеток коры принимают информацию от любых рецепторов организма – это полисенсорные нейроны , воспринимающие импульсы только от определенных рецепторов (зрительных, слуховых, тактильных и т.д.). Клетки нейроглии выполняют вспомогательные функции: трофическую, нейросекреторную, защитную, изолирующую.
Специализированные нейроны и другие клетки, входящие в состав вертикальных колонок, образуют отдельные участки коры, которые называютсяпроекционными зонами (цитоархитектоническими полями) 1 . Эти функциональные зоны коры делятся на 3 группы :
– афферентные (чувственные);
– эфферентные (двигательные или моторные);
– ассоциативные (соединяют предыдущие зоны и обусловливают сложную работу мозга, лежащую в основе высшей психической деятельности).
У человека ассоциативные зоны достигают наибольшего развития. Локализация функций в коре головного мозга относительна – здесь нельзя провести каких-либо четких границ, поэтому мозг обладает высокой пластичностью, приспосабливаемостью к повреждениям. Тем не менее, морфологическая и функциональная неоднородность коры позволила выделить в ней 52 цитоархитектонических поля (К. Бродман), а среди них – центры зрения, слуха, осязания и др. Все они связаны между собой волокнами проводящих путей белого вещества, которые делятся на 3 типа :
1 – ассоциативные (связывают зоны коры в пределах одного полушария),
2 – комиссуральные (связывают симметричные зоны коры двух полушарий через мозолистое тело),
3 – проекционные (связывают кору и подкорку с периферическими органами, бывают чувствительные и двигательные).
Значение важнейших зон коры головного мозга.
1. Чувствительная зона коры (в постцентральной извилине) воспринимает импульсы от тактильных, температурных и болевых рецепторов кожи, а также от проприорецепторов противоположной половины тела.
2. Двигательная зона коры (в предцентральной извилине) содержит в 5 слое коры пирамидные клетки Беца , от которых идут импульсы произвольных движений к скелетным мышцам противоположной половины тела.
3. Премоторная зона (в основании средней лобной извилины) обеспечивает сочетанный поворот головы и глаз в противоположную сторону.
4. Праксическая зона (в надкраевой извилине) обеспечивает сложные целенаправленные движения практической деятельности и профессиональных двигательных навыков. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
5. Центр проприоцептивного гнозиса (в верхней теменной дольке) обеспечивает восприятие импульсов проприорецепторов, контролирует ощущения тела и его частей как целостного образования.
6. Центр чтения (в верхней теменной дольке, вблизи затылочной доли) контролирует восприятие написанного текста.
7. Слуховая зона коры (в верхней височной извилине) воспринимает информацию от рецепторов органа слуха.
8. Слуховой центр речи, центр Вернике (в основании верхней височной извилины). Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
9. Слуховой центр пения (в верхней височной извилине). Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
10. Двигательный центр устной речи, центр Брока (в основании нижней лобной извилины) контролирует произвольные сокращения мышц, участвующих в речеобразовании. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
11. Двигательный центр письменной речи (в основании средней лобной извилины) обеспечивает произвольные движения, связанные с написанием букв и других знаков. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
12. Стереогностическая зона (в угловой извилине) контролирует узнавание предметов наощупь (стереогноз).
13. Зрительная зона коры (в затылочной доле) воспринимает информацию от рецепторов органа зрения.
14. Зрительный центр речи (в угловой извилине) контролирует движение губ и мимику говорящего оппонента, тесно связан с другими сенсорными и моторными речевыми центрами. Речь и сознание – это филогенетические наиболее молодые функции мозга, поэтому речевые центры имеют большое число рассеянных элементов и наименее локализованы. Речевые и мыслительные функции выполняются при участии всей коры. Речевые центры у человека сформировались на основе трудовой деятельности, поэтому они асимметричные, непарные и связаны с рабочей рукой.
При поражении чувствительной зоны коры может возникать частичная потеря чувствительности (гипэстезия ). Одностороннее поражение приводит к нарушению кожной чувствительности на противоположной стороне тела. При двустороннем повреждении наблюдается полная потеря чувствительности (анестезия ). В зависимости от обширности поражения двигательной зоны коры возникает частичная (парез ) или полная (паралич ) утрата движений. При поражении праксической зоны развивается (моторная или конструктивная ) апраксия. Апраксия другого рода (идеаторная апраксия – «апраксия замысла» ) возникает при поражении передних отделов лобных долей. Здесь же возможно нарушение координации движений (корковая атаксия ), сложных двигательных функций (акинезия ), обеспечивающих трудовую деятельность, письмо (аграфия ) и речь (моторная афазия ). Поражение центра проприоцептивного гнозиса вызывает агнозию частей собственного тела (аутотопагнозию ) – нарушение схемы тела. Поражение стереогностической зоны приводит к потере способности чтения (алексия ). При двустороннем поражении слуховой зоны коры возникает полная корковая глухота. Поражение слухового центра речи (Вернике) имеет место словесная глухота (сенсорная афазия ), а при поражении слухового центра пения возникает музыкальная глухота (сенсорная амузия) и неспособность к составлению осмысленных предложений из отдельных слов (аграмматизм ). Поражение зрительной зоны коры в равных ее участках вызывает утрату способности ориентироваться в незнакомой обстановке, потерю зрительной памяти. Двустороннее поражение приводит к полной корковой слепоте.
Любая функциональная зона коры находится в анатомической и функциональной связи с другими зонами коры, с подкорковыми ядрами, структурами промежуточного мозга и ретикулярной формации, что обеспечивает совершенство выполняемых ими функций.
Лимбическая система – наиболее древняя часть коры, включающая в себя ряд образований коркового и подкоркового уровня (лобные доли мозга, поясная извилина, мозолистое тело, серый покров, свод, гиппокамп, миндалина и сосцевидные тела, таламус, стриопаллидарная система, ретикулярная формация). Основные ее функции :
1 – регуляция вегетативных процессов (особенно пищеварения),
2 – регуляция поведенческих реакций,
3 – формирование и регуляция эмоций, сна,
4 – формирование и проявление памяти.
Лимбическая система формирует положительные и отрицательные эмоции со всеми сопровождающими и вегетативными, эндокринными и двигательными компонентами 2 . Она создает мотивацию поведения , просчитывает способы действий, пути достижения полезного результата. Способность воссоздавать перед глазами прошедшие события – одна из удивительных способностей мозга. Ключевая роль в обработке информации принадлежит гиппокампу (морской конек). Здесь происходит ее качественная сортировка. Часть информации попадает в ассоциативные зоны коры и там анализируется, а другая часть сразу закрепляется в долговременной памяти. Отдельные воспоминания систематизируются и превращаются в устойчивые во сне, в фазе глубокого сна, когда человек не видит сновидений.
При поражении лимбической системы затрудняется формирование условных рефлексов, нарушаются процессы памяти, теряется избирательность реакций и отмечается неумеренное их усиление.
Большой мозг состоит из почти идентичных половин – правого и левого полушарий, которые связаны мозолистым телом. Комиссуральные волокна связывают симметричные зоны коры. Тем не менее, кора правого и левого полушарий не симметричны не только внешне, но и в функциональном отношении. Установлено, что левое полушарие обеспечивает логическое абстрактное мышление . Оно отвечает за письмо, чтение, математический счет. Правое полушарие обеспечивает конкретное образное мышление . Оно отвечает за эмоциональную окраску речи, музыкальность, ориентацию в пространстве, восприятие геометрических фигур, рисунков, природных объектов.
Оба полушария работают вместе, но одно из них, как правило, доминирует у каждого человека. По способу мышления и характеру запоминания информации все люди практически делятся на левополушарный тип и правополушарный тип 3 . Темпы созревания левого и правого полушария имеют половые особенности. У девочек левое полушарие развивается быстрее, что подтверждается более быстрым развитием речи и психомоторным развитием. У аномальных детей развитие левого полушария значительно задерживается, функциональная асимметрия выражена слабо. У детей с высокой умственной работоспособностью сильнее выражено различие между правым и левым полушарием (Физиол. человека, № 1, 1983 г.).
Для изучения функций коры головного мозга применяются различные методы :
1. Удаление отдельных участков коры оперативным путем (экстирпация).
2. Метод раздражения электрическими, химическими и температурными раздражителями.
3. Метод отведения биопотенциалов и регистрации электрической активности зон коры или отдельных нейронов, ЭЭГ.
4. Классический метод условных рефлексов.
5. Клинический метод изучения функций у людей с поражениями коры мозга.
6. Техника сканирования, например ядерно-магнитный резонанс и позитрон-эмиссионная томография. Пользуясь этими методами, наблюдая за притоком крови к определенным областям мозга во время мыслительных процессов, исследователи установили, какие именно участки коры помогают слышать слова, видеть слова и произносить слова.
7. Метод тепловизионного исследования позволил уточнить гипотезу о том, что, несмотря на сложную структуру коры, можно увидеть изображение на ее поверхности. Эту гипотезу выдвинули ученые Института ВНД и нейрофизиологии. Сотрудники Института радиотехники и электроники АН РФ гипотезу подтвердили. Тепловизор с чувствительностью в сотые доли градуса передавал в компьютер термокарты коры головного мозга белой крысы со скоростью 25 кадров в секунду. Крысе показывали изображения геометрических фигур. На дисплее эти фигуры четко просматривались на поверхности коры мозга. Первичное изображение, попадающее на сетчатку, преобразуется рецепторами в импульсы и вновь восстанавливается в коре как на экране.
Электроэнцефалография (ЭЭГ) является распространенным методом исследования мозга. Ритм электрических колебаний соответствует тому или иному функциональному состоянию мозга.
Активное бодрствование сопровождает (бета)-ритм с частотой 14-100 колебаний в секунду.
В покое с закрытыми глазами наблюдается (альфа)- ритм с частотой 8 – 13 колебаний в секунду.
Во время глубокого сна регистрируется (дельта)- ритм с частотой 0,5-3 колебаний в секунду.
В состоянии неглубокого сна наблюдается (тета) - ритм с частотой 4-7 колебаний в секунду.
Ээг позволяет объективно оценить подвижность, распространенность и взаимоотношения в коре процессов возбуждения и торможения.
ОПРЕДЕЛЕНИЕ
Анализатор - функциональная единица, отвечающая за восприятие и анализ сенсорной информации одного вида (термин ввел И. П. Павлов).
Анализатор представляет собой совокупность нейронов, участвующих в восприятии раздражений, проведении возбуждения и в анализе раздражения.
Анализатор часто называют сенсорной системой . Анализаторы классифицируют по типу тех ощущений, в формировании которых они участвуют (см. рис. ниже).
Рис. Анализаторы
Это зрительный, слуховой, вестибулярный, вкусовой, обонятельный, кожный, мышечный и другие анализаторы. В анализаторе выделяют три отдела:
- Периферический отдел : рецептор, предназначенный для преобразования энергии раздражения в процесс нервного возбуждения.
- Проводниковый отдел : цепь из центростремительных (афферентных) и вставочных нейронов, по которой импульсы передаются от рецепторов к вышележащим отделам центральной нервной системы.
- Центральный отдел : определенная зона коры больших полушарий.
Кроме восходящих (афферентных) путей существуют нисходящие волокна (эфферентные), по которым осуществляется регуляция деятельности нижних уровней анализатора со стороны его высших, в особенности корковых, отделов.
| анализатор | периферический отдел (орган чувств и рецепторы) | проводниковый отдел | центральный отдел |
|---|---|---|---|
| зрительный | рецепторы сетчатки глаза | зрительный нерв | зрительный центр в затылочной доле КБП |
| слуховой | чувствительные волосковые клетки кортиева (спирального) органа улитки | слуховой нерв | слуховой центр в височной доле КБП |
| обонятельный | обонятельные рецепторы эпителия носа | обонятельный нерв | обонятельный центр в височной доле КБП |
| вкусовой | вкусовые почки ротовой полости (в основном, корня языка) | языкоглоточный нерв | вкусовой центр в височной доле КБП |
| осязательный (тактильный) | осязательные тельца сосочкового слоя дермы (болевые, температурные, тактильные и др. рецепторы) | центростремительные нервы; спинной, продолговатый, промежуточный мозг | центр кожной чувствительности в центральной извилине теменной доли КБП |
| кожно-мышечный | проприорецепторы в мышцах и связках | центростремительные нервы; спинной мозг;продолговатый и промежуточный мозг | двигательная зона и прилегающим к ней участки лобной и теменных долей. |
| вестибулярный | полукружные канальца и преддверие внутреннего уха | преддверно-улитковый нерв (VIII пара черепно-мозговых нервов) | мозжечок |
КБП* - кора больших полушарий.
органы чувств
Человек обладает рядом важных специализированных периферических образований -органов чувств , обеспечивающих восприятие воздействующих на организм внешних раздражителей.
Орган чувств состоит из рецепторов и вспомогательного аппарата, который помогает улавливать, концентрировать, фокусировать, направлять и т. д. сигнал.
К органам чувств относятся органы зрения, слуха, обоняния, вкуса, осязания. Сами по себе они не могут обеспечить ощущение. Для возникновения субъективного ощущения необходимо, чтобы возбуждение, возникшее в рецепторах, поступило в соответствующий отдел коры больших полушарий.
Структурные поля коры больших полушарий
Если рассматривать стуктурную организацию коры больших полушарий, то можно выделить несколько полей, имеющих различное клеточное строение.
Различают три основные группы полей в коре:
- первичные
- вторичные
- третичные.
Первичные поля , или ядерные зоны анализаторов, непосредственно связаны с органами чувств и органами движения.
Например, поле болевой, температурной, кожно-мышечной чувствительности в задней части центральной извилины, зрительное поле в затылочной доле, слуховое поле в височной доле и двигательное поле в передней части центральной извилины.
Первичные поля они раньше других созревают в онтогенезе.
Функция первичных полей: анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов.
При разрушении первичных полей возникает так называемая корковая слепота, корковая глухота и т. п.
Вторичные поля расположены рядом с первичными и связаны через них с органами чувств.
Функция вторичных полей: обобщение и дальнейшая обработка поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия.
При поражении вторичных полей человек видит и слышит, но не способен осознать, понять значение увиденного и услышанного.
Первичные и вторичные поля имеются и у человека, и у животных.
Третичные поля , или зоны перекрытия анализаторов, находятся в задней половине коры - на границе теменной, височных и затылочной долей и в передних частях лобных долей. Они занимают половину всей площади коры больших полушарий и имеют многочисленные связи со всеми ее частями. В третичных полях оканчивается большинство нервных волокон, соединяющих левое и правое полушария.
Функция третичных полей: организация согласованной работы обоих полушарий, анализ всех воспринятых сигналов, их сравнение с ранее полученнойнформацией, координация соответствующего поведения, программирование двигательной активности.
Эти поля есть только у человекаи созревают позже других корковых полей.
Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью и даже простейшими двигательными навыками.

Рис. Структурные поля коры больших полушарий
С учетом расположения структурных полей коры больших полушарий можно выделить функциональные части: сенсорные, моторные и ассоциативные зоны.
Все сенсорные и моторные зоны занимают менее 20% поверхности коры. Остальная кора составляет ассоциативную область.
Ассоциативные зоны
Ассоциативные зоны - это функциональные зоны коры головного мозга. Они связывают вновь поступающую сенсорную информацию с полученной ранее и хранящейся в блоках памяти, а также сравнивают между собой информацию, получаемую от разных рецепторов (см. рис. ниже).
Каждая ассоциативная область коры связана с несколькими структурными полями. В состав ассоциативных зон входит часть теменной, лобной и височной долей. Границы ассоциативных зон нечеткие, ее нейроны участвуют в интеграции различной информации. Здесь идет высший анализ и синтез раздражений. В результате формируются сложные элементы сознания.

Рис. Борозды и доли коры больших полушарий
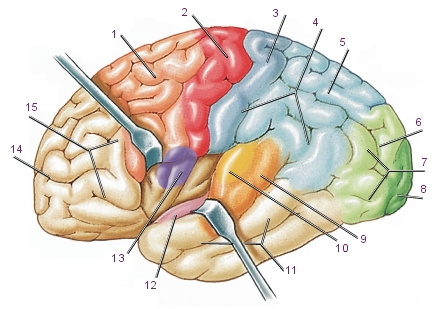
Рис. Ассоциативные зоны коры больших полушарий:
1. Ассоциативная двигател ьная зона (лобная доля)
2. Первичная двигательная зона
3. Первичная соматосенсорная зона
4. Теменная доля больших полушарий
5. Ассоциативная соматосенсорная (кожно-мышечная) зона (теменная доля)
6. Ассоциативная зрительная зона (затылочная доля)
7. Затылочная доля больших полушарий
8. Первичная зрительная зона
9. Ассоциативная слуховая зона (височные доли)
10. Первичная слуховая зона
11. Височная доля больших полушарий
12. Обонятельная кора (внутренняя поверхность височной доли)
13. Вкусовая кора
14. Предлобная ассоциативная зона
15. Лобная доля больших полушарий.
Сенсорные сигналы в ассоциативной зоне расшифровываются, осмысливаются и используются для определения наиболее подходящих ответных реакций, которые передаются в связанную с ней двигательную (моторную) зону.
Таким образом, ассоциативные зоны участвуют в процессах запоминания, обучения и мышления, и результаты их деятельности составляют интеллект (способность организма использовать полученные знания).
Отдельные крупные ассоциативные области расположены в коре рядом с соответствующими сенсорными зонами. Например, зрительная ассоциативная зона расположена в затылочной зоне непосредственно впереди сенсорной зрительной зоны и осуществляет полную обработку зрительной информации.
Некоторые ассоциативные зоны выполняют только часть обработки информации и связаны с другим ассоциативными центрами, выполняющими дальнейшую обработку. Например, звуковая ассоциативная зона анализирует звуки, разделяя их на категории, а затем передает сигналы в более специализированные зоны, такие как речевая ассоциативная зона, где воспринимается смысл услышанных слов.
Эти зоны относятся к ассоциативной коре и участвуют в организации сложных форм поведения.
В коре больших полушарий выделяют области с менее определенными функциями. Так, значительная часть лобных долей, особенно с правой стороны, может быть удалена без заметных нарушений. Однако, если произвести двухстороннее удаление лобных областей возникают тяжелые психические нарушения.
вкусовой анализатор
Вкусовой анализатор отвечает за восприятие и анализ вкусовых ощущений.
Периферический отдел : рецепторы - вкусовые луковицы в слизистой оболочке языка, мягкого неба, миндалин и других органов ротовой полости.


Рис. 1. Вкусовой сосочек и вкусовая луковица
Вкусовые сосочки несут на боковой поверхности вкусовые луковицы (рис. 1, 2), в состав которых входят 30 - 80 чувствительных клеток. Вкусовые клетки усеяны на своем конце микроворсинками - вкусовыми волосками. Они выходят на поверхность языка через вкусовые поры. Вкусовые клетки непрерывно делятся и непрерывно гибнут. Особенно быстро происходит замещение клеток, расположенных в передней части языка, где они лежат более поверхностно.

Рис. 2. Вкусовая луковица: 1 - нервные вкусовые волокна; 2 - вкусовая почка (чашечка); 3 - вкусовые клетки; 4 - поддерживающие (опорные) клетки; 5 - вкусовая пора

Рис. 3. Вкусовые зоны языка: сладкое - кончик языка; горькое - основание языка; кислое - боковая поверхность языка; соленое - кончик языка.
Вкусовые ощущения вызывают только растворенные в воде вещества.
Проводниковый отдел : волокна лицевого и языкоглоточного нерва (рис. 4).
Центральный отдел : внутренняя сторона височной доли коры больших полушарий.

обонятельный анализатор
Обонятельный анализатор отвечает за восприятие и анализ запаха.
- пищевое поведение;
- апробация пищи на съедобность;
- настройка пищеварительного аппарата на обработку пищи (по механизму условного рефлекса);
- оборонительное поведение (в т. ч. проявление агрессии).
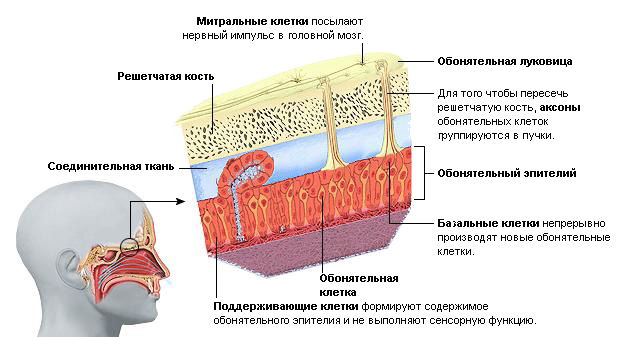
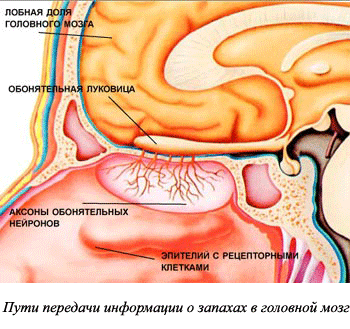
Периферический отдел: рецепторы слизистой оболочки верхней части носовой полости. Обонятельные рецепторы в слизистой носа оканчиваются обонятельными ресничками. Газообразные вещества растворяются в слизи, окружающей реснички, затем в результате химической реакции возникает нервный импульс (рис. 5).
Проводниковый отдел: обонятельный нерв.
Центральный отдел : обонятельная луковица (структура переднего мозга, в которой осуществляется обработка информации) и обонятельный центр, расположенный на нижней поверхности височной и лобной долей коры больших полушарий (рис. 6).
В коре происходит определение запаха и формируется адекватная на него реакция организма.
Восприятие вкуса и запаха дополняют друг друга, давая целостное представление о виде и качестве пищи. Оба анализатора связаны с центром слюноотделения продолговатого мозга и участвуют в пищевых реакциях организма.
Осязательный и мышечный анализатор объединяют в соматосенсорную систему - систему кожно-мышечной чувствительности.
Строение соматосенсорного анализатора
Периферический отдел : проприорецепторы мышц и сухожилий; рецепторы кожи (механорецепторы, терморецепторы и др.).
Проводниковый отдел : афферентные (чувствительны) нейроны; восходящие пути спинного мозга; продолговатый мозг, ядра промежуточного мозга.
Центральный отдел : сенсорная зона в теменной доле коры больших полушарий.
Рецепторы кожи
Кожа является самым крупным чувствительный органом в теле человека. На ее поверхности (около 2 м2) сосредоточено множество рецепторов.
Большинство ученых склоняются к наличию четырех основных видов кожной чувствительности: тактильной, тепловой, холодовой и болевой.
Рецепторы распределены неравномерно и на разной глубине. Больше всего рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов.
МЕХАНОРЕЦЕПТОРЫ КОЖИ
- тонкие окончания нервных волокон , оплетающие кровеносные сосуды, волосяные сумки и т.п.
- клетки Меркеля - нервные окончания базального слоя эпидермиса (много на подушечках пальцев);
- осязательные тельца Мейсснера - сложные рецепторы сосочкового слоя дермы (много на пальцах, ладонях, подошвах, губах, языке, половых органах и сосках молочных желез);
- пластинчатые тельца - рецепторы давления и вибрации; расположены в глубоких слоях кожи, в сухожилиях, связках и брыжейке;
- луковицы (колбы Краузе) - нервные рецепторы в соединительнотканном слое слизистых оболочек, под эпидермисом и среди мышечных волокон языка.
МЕХАНИЗМ РАБОТЫ МЕХАНОРЕЦЕПТОРОВ
Механический стимул - деформация мембраны рецептора - уменьшение электрического сопротивления мембраны - увеличение проницаемости мембраны для Na+ - деполяризация мембраны рецептора - распространение нервного импульса
АДАПТАЦИЯ КОЖНЫХ МЕХАНОРЕЦЕПТОРОВ
- быстро адаптирующиеся рецепторы : кожные механорецепторы в волосяных луковицах, пластинчатые тельца (не ощущаем давление одежды, контактных линз и т.п.);
- медленно адаптирующиеся рецепторы: осязательные тельца Мейсснера.
Ощущение прикосновения и давления на кожу довольно точно локализуется, т. е. относится человеком к определенному участку кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенезе при участии зрения и проприорецепции.
Способность человека раздельно воспринимать прикосновение к двум соседним точкам кожи, также сильно отличается в разных ее участках. На слизистой оболочке языка порог пространственного различия равен 0,5 мм, а на коже спины - более 60 мм.
Температурная рецепция
Температура тела человека колеблется в сравнительно узких пределах, поэтому информация о температуре окружающей среды, необходимая для деятельности механизмов терморегуляции, имеет особо важное значение.
Терморецепторы располагаются в коже, роговице глаза, в слизистых оболочках, а также в ЦНС (в гипоталамусе).
ВИДЫ ТЕРМОРЕЦЕПТОРОВ
- холодовые терморецепторы : многочисленные; лежат близко к поверхности.
- тепловые терморецепторы : их значительно меньше; лежат в более глубоком слое кожи.
- специфические терморецепторы : воспринимают только температуру;
- неспецифические терморецепторы : воспринимают температурные и механические раздражители.
Терморецепторы реагируют на изменение температуры повышением частоты генерируемых импульсов, устойчиво длящимся все время действия стимула. Изменение температуры на 0,2 °С вызывает длительные изменения их импульсации.
В некоторых условиях холодовые рецепторы могут быть возбуждены теплом, а тепловые холодом. Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну или обжигающее действие ледяной воды.
Начальные температурные ощущения зависят от разницы температуры кожи и температуры действующего раздражителя, его площади и места приложения. Так, если руку держали в воде температуры 27 °С, то в первый момент при переносе руки в воду, нагретую до 25 °С, она кажется холодной, однако уже через несколько секунд становится возможной истинная оценка абсолютной температуры воды.
Болевая рецепция
Болевая чувствительность имеет первостепенное значение для выживания организма, являясь сигналом об опасности при сильных воздействиях различных факторов.
Импульсы болевых рецепторов часто свидетельствуют о патологических процессах в организме.
На данный момент не найдены специфическе болевые рецепторы.
Сформулированы две гипотезы об организации болевого восприятия:
- Существуют специфические болевые рецепторы - свободные нервные окончания с высоким порогом реакции;
- Специфических болевых рецепторов не существует; боль возникает при сверхсильном раздражении любых рецепторов.
Механизм возбуждения рецепторов при болевых воздействиях пока не выяснен.
Наиболее общей причиной возникновения боли можно считать изменение концентрации Н+ при токсическом воздействии на дыхательные ферменты или при повреждении клеточных мембран.
Одной из возможных причин длительной жгучей боли может быть выделение при повреждении клеток гистамина, протеолитических ферментов и др. веществ, вызывающих цепочку биохимических реакций, приводящих к возбуждению нервных окончаний.
Болевая чувствительность практически не представлена на корковом уровне, поэтому высшим центром болевой чувствительности является таламус, где 60 % нейронов в соответствующих ядрах.четко реагирует на болевое раздражение.
АДАПТАЦИЯ БОЛЕВЫХ РЕЦЕПТОРОВ
Адаптация болевых рецепторов зависит от многочисленных факторов и ее механизмы мало изучены.
Например, заноза, будучи неподвижной, не вызывает особых болевых ощущений. Пожилые люди в некоторых случаях "привыкают не замечать" головной боли или боли в суставах.
Однако в очень многих случаях болевые рецепторы не обнаруживают существенной адаптации, что делает страдания больного особенно длительными и мучительными и требует применения анальгетиков.
Болевые раздражения вызывают ряд рефлекторных соматических и вегетативных реакций. При умеренной выраженности эти реакции имеют приспособительное значение, но могут привести к тяжелым патологическим эффектам, например к шоку. Среди этих реакций отмечают повышение мышечного тонуса, частоты сердечных сокращений и дыхания, повышение ил понижение давления, сужение зрачков, увеличение содержания глюкозы в крови и ряд других эффектов.
ЛОКАЛИЗАЦИЯ БОЛЕВОЙ ЧУВСТВИТЕЛЬНОСТИ
При болевых воздействиях на кожу человек локализует их достаточно точно, но при заболеваниях внутренних органов могут вознкать отраженные боли . Например, при почечной колике, больные жалуются на "вступающие" резкие боли в ногах и прямой кишке. Могут быть и обратные эффекты.
проприорецепция
Виды проприорецепторов:
- нервно-мышечные веретена: дают информацию о скорости и силе мышечного растяжения и сокращения;
- сухожильные рецепторы Гольджи: дают информацию о силе мышечного сокращения.
Функции проприорецепторов:
- восприятие механических раздражений;
- восприятие пространственного расположения частей тела.
НЕРВНО-МЫШЕЧНОЕ ВЕРЕТЕНО
Нервно-мышечное веретено - сложный рецептор, который включает видоизмененные мышечные клетки, афферентные и эфферентные нервные отростки и контролирует как скорость, так и степень сокращения и растяжение скелетных мышц.
Нервно-мышечное веретено расположено в толще мышцы. Каждое веретено покрыто капсулой. Внутри капсулы находится пучок специальных мышечных волокон. Веретена расположены параллельно волокнам скелетных мышц, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении - уменьшается.

Рис. Нервно-мышечное веретено
СУХОЖИЛЬНЫЕ РЕЦЕПТОРЫ ГОЛЬДЖИ
Находятся в зоне соединения мышечных волокон с сухожилием.
Сухожильные рецепторы слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении. Интенсивность их импульсации примерно пропорциональна силе сокращения мышцы.

Рис. Сухожильный рецептор Гольджи
СУСТАВНЫЕ РЕЦЕПТОРЫ
Они изучены меньше, чем мышечные. Известно, что суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя таким образом в системе обратных связей от двигательного аппарата и в управлении им.
Зрительный анализатор включает:
- периферический отдел: рецепторы сетчатки глаза;
- проводниковый отдел: зрительный нерв;
- центральный отдел: затылочная доля коры больших полушарий.
Функция зрительного анализатора : восприятие, проведение и расшифровка зрительных сигналов.
Строения глаза
Глаз состоит из глазного яблока и вспомогательного аппарата .
Вспомогательный аппарат глаза
- брови - защита от пота;
- ресницы - защита от пыли;
- веки - механическая защита и поддержание влажности;
- слезные железы - расположены у верхней части наружного края глазницы. Она выделяет слезную жидкость, увлажняющую, промывающую и дезинфицирующую глаз. Избыток слёзной жидкости удаляется в носовую полость через слёзный канал , расположенный во внутреннем углу глазницы.
ГЛАЗНОЕ ЯБЛОКО
Глазное яблоко имеет примерно сферическую форму с диаметром около 2,5 см.
Оно расположено на жировой подушке в переднем отделе глазницы.
Глаз имеет три оболочки:
- белочная оболочка (склера) с прозрачной роговицей - наружная очень плотная фиброзная оболочка глаза;
- сосудистая оболочка с наружной радужной оболочкой и ресничным телом - пронизана кровеносными сосудами (питание глаза) и содержит пигмент, препятствующий рассеиванию света через склеру;
- сетчатая оболочка (сетчатка ) - внутренняя оболочка глазного яблока - рецепторная часть зрительного анализатора; функция: непосредственное восприятие света и передача информации в центральную нервную систему.

Коньюктива - слизистая оболочка, соединяющая глазное яблоко с кожным покровами.
Белочная оболочка (склера) - внешняя прочная оболочка глаза; внутренняя часть склеры непроницаема для сетовых лучей. Функция: защита глаза от внешних воздействий и светоизоляция;
Роговица - передняя прозрачная часть склеры; является первой линзой на пути световых лучей. Функция: механическая защита глаза и пропускание световых лучей.
Хрусталик - двояковыпуклая линза, расположенная за роговицей. Функция хрусталика: фокусировка световых лучей. Хрусталик не имеет сосудов и нервов. В нем не развиваются воспалительные процессы. В нем много белков, которые иногда могут терять свою прозрачность, что приводит к заболеванию, называемому катаракта .
Сосудистая оболочка - средняя оболочка глаза, богатая сосудами и пигментом.
Радужная оболочка - передняя пигментированная часть сосудистой оболочки; содержит пигменты меланин и липофусцин, определяющие цвет глаз.
Зрачок - круглое отверстие в радужной оболочке. Функция: регуляция светового потока, поступающего в глаз. Диаметр зрачка непроизвольно меняется с помощью гладких мышц радужной оболочки при изменении освещенности.
Передняя и задняя камеры - пространство спереди и сзади радужной оболочки, заполненное прозрачной жидкостью (водянистой влагой ).
Ресничное (цилиарное) тело - часть средней (сосудистой) оболочки глаза; функция: фиксация хрусталика, обеспечение процесса аккомодации (изменение кривизны) хрусталика; продуцирование водянистой влаги камер глаза, терморегуляция.
Стекловидное тело - полость глаза между хрусталиком и глазным дном , заполненная прозрачным вязким гелем, поддерживающим форму глаза.
Сетчатка (ретина) - рецепторный аппарат глаза.
СТРОЕНИЕ СЕТЧАТКИ
Сетчатка образована разветвлениями окончаний зрительного нерва, который, подойдя к глазному яблоку, проходит через белочную оболочку, причем оболочка нерва сливается с белочной оболочкой глаза. Внутри глаза волокна нерва распределяются в виде тонкой сетчатой оболочки, которая выстилает задние 2/3 внутренней поверхности глазного яблока.
Сетчатка состоит из опорных клеток, образующих сетчатую структуру, откуда и произошло ее название. Световые лучи воспринимает только ее задняя часть. Сетчатая оболочка по своему развитию и по функции представляет собой часть нервной системы. Все же остальные части глазного яблока играют вспомогательную роль для восприятия сетчаткой зрительных раздражений.
Сетчатая оболочка - это часть мозга, выдвинутая наружу, ближе к поверхности тела, и сохраняющая с ним связь с помощью пары зрительных нервов.
Нервные клетки образуют в сетчатке цепи, состоящие из трех нейронов (см. рис. ниже):
- первые нейроны имеют дендриты в виде палочек и колбочек; эти нейроны являются конечными клетками зрительного нерва, они воспринимают зрительные раздражения и представляют собой световые рецепторы.
- вторые - биполярные нейроны;
- третьи - мультиполярные нейроны (ганглиозные клетки ); от них отходят аксоны, которые тянутся по дну глаза и образуют зрительный нерв.

Светочувствительные элементы сетчатки:
- палочки - воспринимают яркость;
- колбочки - воспринимают цвет.
Колбочки медленно возбуждаются и только ярким светом. Они способны воспринимать цвет. В сетчатке находится три вида колбочек. Первые воспринимают красный цвет, вторые - зеленый, третьи - синий. В зависимости от степени возбуждения колбочек и сочетания раздражений, глаз воспринимает различные цвета и оттенки.
Палочки и колбочки в сетчатой оболочке глаза перемешаны между собой, но в некоторых местах они расположены очень густо, в других же редко или отсутствуют совсем. На каждое нервное волокно приходится примерно 8 колбочек и около 130 палочек.
В области желтого пятна на сетчатке нет палочек - только колбочки, здесь глаз обладает наибольшей остротой зрения и наилучшим восприятием цвета. По-этому глазное яблоко находится в непрерывном движении, так чтобы рассматриваемая часть объекта приходилась на желтое пятно. По мере удаления от желтого пятна плотность палочек увеличивается, но потом уменьшается.
При низкой освещенности в процессе видения участвуют только палочки (сумеречное видение), и глаз не различает цвета, зрение оказывается ахроматическим (бесцветным).
От палочек и колбочек отходят нервные волокна, которые, соединяясь, образуют зрительный нерв. Место выхода из сетчатки зрительного нерва называется диском зрительного нерва . В области диска зрительного нерва светочувствительных элементов нет. Поэтому это место не дает зрительного ощущения и называется слепым пятном .
МЫШЦЫ ГЛАЗА
- глазодвигательные мышцы - три пары поперечно-полосатых скелетных мышц, которые прикрепляются к коньюктиве; осуществляют движение глазного яблока;
- мышцы зрачка
- гладкие мышцы радужки (круговая и радиальная), меняющие диаметр зрачка;
Круговая мышца (сжиматель) зрачка иннервируется парасимпатическими волокнами из глазодвигательного нерва, а радиальная мышца (расширитель) зрачка - волокнами симпатического нерва. Радужная оболочка, таким образом, регулирует количество света, поступающего в глаз; при сильном, ярком свете зрачок суживается и ограничивает поступление лучей, а при слабом - расширяется, давая возможность проникнуть большему количеству лучей. На диаметр зрачка влияет гормон адреналин. Когда человек находится в возбужденном состоянии (при испуге, гневе и т. д.), количество адреналина в крови увеличивается, и это вызывает расширение зрачка.
Движения мышц обоих зрачков управляются из одного центра и происходят синхронно. Поэтому оба зрачка всегда одинаково расширяются или суживаются. Даже если подействовать ярким светом на один только глаз, зрачок другого глаза тоже суживается. - мышцы хрусталика (цилиарные мышцы) - гладкие мышцы, изменяющие кривизну хрусталика (аккомодация --фокусировка изображения на сетчатке).
Проводниковый отдел
Зрительный нерв является проводником световых раздражений от глаза к зрительному центру и содержит чувствительные волокна.
Отойдя от заднего полюса глазного яблока, зрительный нерв выходит из глазницы и, войдя в полость черепа, через зрительный канал, вместе с таким же нервом другой стороны, образует перекрест (хиазму ) под гиполаламусом. После перекреста зрительные нервы продолжаются в зрительных трактах . Зрительный нерв связан с ядрами промежуточного мозга, а через них - с корой больших полушарий.
Каждый зрительный нерв содержит совокупность всех отростков нервных клеток сетчатки одного глаза. В области хиазмы происходит неполный перекрест волокон, и в составе каждого зрительного тракта оказывается около 50% волокон противоположной стороны и столько же волокон своей стороны.
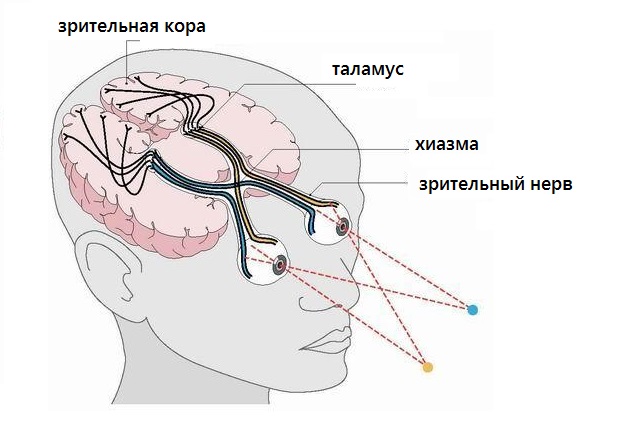
Центральный отдел
Центральный отдел зрительного анализатора расположен в затылочной доле коры больших полушарий.
Импульсы от световых раздражений по зрительному нерву проходят к мозговой коре затылочной доли, где расположен зрительный центр.
В волокна каждого нерва связаны с двумя полушариями мозга, причем изображение, получаемое на левой половине сетчатки каждого глаза, анализируется в зрительной коре левого полушария, а на правой половине сетчатки - в коре правого полушария.
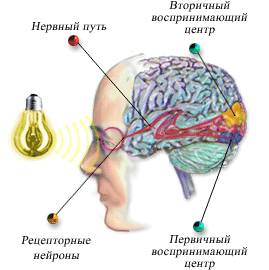
нарушение зрения
С возрастом и под воздействием других причин способность управлять кривизной поверхности хрусталика ослабевает.
Близорукость (миопия) - фокусировка изображение перед сетчаткой; развивается из-за увеличения кривизны хрусталика, которая может возникнуть при неправильном обмене веществ или нарушении гигиены зрения. И справляют очками с вогнутыми линзами.
Дальнозоркость - фокусировка изображения позади сетчатки; возникает вследствие уменьшения выпуклости хрусталика. И справляют очками с выпуклыми линзами.

Существует два пути проведения звуков:
- воздушная проводимость : через наружный слуховой проход, барабанную перепонку и цепь слуховых косточек;
- тканевая проводимост ь: через ткани черепа.
Функция слухового анализатора: восприятие и анализ звуковых раздражений.
Периферический отдел: слуховые рецепторы в полости внутреннего уха.
Проводниковый отдел: слуховой нерв.
Центральный отдел: слуховая зона в височной доле коры больших полушарий.


Рис. Височная кость Рис. Расположение органа слуха в полости височной кости
строение уха
Орган слуха у человека расположен в полости черепа в толще височной кости.
Он делится на три отдела: наружное, среднее и внутреннее ухо. Эти отделы тесно связаны анатомически и функционально.
Наружное ухо состоит из наружного слухового прохода и ушной раковины.
Среднее ухо - барабанная полость; она отделена барабанной перепонкой от наружного уха.
Внутреннее ухо, или лабиринт , - отдел уха, где происходит раздражение рецепторов слухового (улиткового) нерва; он помещается внутри пирамиды височной кости. Внутреннее ухо образует орган слуха и равновесия.
Наружное и среднее ухо имеют второстепенное значение: они проводят звуковые колебания к внутреннему уху, и таким образом является звукопроводящим аппаратом.

Рис. Отделы уха
НАРУЖНОЕ УХО
Наружное ухо включает ушную раковину и наружный слуховой проход , которые предназначены для улавливания и проведения звуковых колебаний.
Ушная раковина образована тремя тканями:
- тонкой пластинкой гиалинового хряща, покрытого с обеих сторон надхрящницей, имеющего сложную выпукло-вогнутую форму, определяющую рельеф ушной раковины;
- кожей очень тонкой, плотно прилегающей к надхрящнице и почти не имеющей жировой клетчатки;
- подкожной жировой клетчаткой, расположенной в значительном количестве в нижнем отделе ушной раковины - мочке уха .
Ушная раковина прикрепляется к височной кости связками и имеет рудиментарные мышцы, которые хорошо выражены у животных.
Ушная раковина устроена так, чтобы максимально концентрировать звуковые колебания и направлять их в наружное слуховое отверстие.
Форма, величина, постановка ушной раковины и размеры ушной дольки индивидуальны у каждого человека.
Дарвинов бугорок - рудиментарный треугольный выступ, который наблюдается у 10% людей в верхне-задней области завитка раковины; он соответствует верхушке уха животных.

Рис. Дарвинов бугорок
Наружный слуховой проход представляет собой S-образную трубку длинной примерно 3 см и диаметром 0,7 см, которая снаружи открывается слуховым отверстием и отделяется от полости среднего уха барабанной перепонкой .
Хрящевая часть, являющаяся продолжением хряща ушной раковины, составляет 1/3 его длины, остальные 2/3 образованы костным каналом височной кости. В месте перехода хрящевого отдела в костный канал сужается и изгибается. В этом месте находится связка из эластичной соединительной ткани. Такое строение делает возможным растяжение хрящевого отдела прохода в длину и в ширину.
В хрящевой части слухового прохода кожа покрыта короткими волосками, предохраняющими от попадания в ухо мелких частиц. В волосяные фолликулы открываются сальные железы. Характерным для кожи этого отдела является наличие в более глубоких слоях серных желез.
Серные железы являются производными потовых желез.Серные железы впадают либо в волосяные фолликулы, либо свободно в кожу. Серные железы выделяют светло-желтый секрет, который вместе с отделяемым сальных желез и с отторгшимся эпителием образует ушную серу .
Ушная сера - светло-желтый секрет серных желез наружного слухового прохода.
Сера состоит из белков, жиров, жирных кислот и минеральных солей. Часть белков являются иммуноглобулинами, определяющими защитную функцию. Кроме того, в состав серы входят отмершие клетки, кожное сало, пыль и другие включения.
Функция ушной серы:
- увлажнение кожи наружного слухового прохода;
- очистки слухового прохода от инородных частиц (пыли, сора, насекомых);
- защита от бактерий, грибков и вирусов;
- жировая смазка в наружной части слухового прохода препятствует попаданию в него воды.
Ушная сера вместе с загрязнениями естественным образом выводится из слухового прохода наружу при жевательных движениях и речи. Кроме этого кожа слухового прохода постоянно обновляется и растет наружу из слухового прохода, вынося с собой серу.
Внутренний костный отдел наружного слухового прохода является каналом височной кости, заканчивающимся барабанной перепонкой. В середине костного отдела расположено сужение слухового прохода - перешеек, за которым расположен более широкий участок.
Кожа костного отдела тонкая, не содержит волосяных луковиц и желез и переходит на барабанную перепонку, образуя ее наружный слой.
Барабанная перепонка представляет собой тонкую овальную (11 x 9 мм) полупрозрачную пластинку, непроницаемую для воды и воздуха. Перепонка состоит из эластических и коллагеновых волокон, которые в верхней ее части замещены волокнами рыхлой соединительной ткани. Со стороны слухового прохода перепонка покрыта плоским эпителием, а со стороны барабанной полости - эпителием слизистой оболочки.
В центральной части барабанная перепонка вогнута, к ней со стороны барабанной полости прикрепляется рукоятка молоточка - первой слуховой косточки среднего уха.
Барабанная перепонка закладывается и развивается вместе с органами наружного уха.
СРЕДНЕЕ УХО
Среднее ухо включает выстланную слизистой оболочкой и заполненную воздухомбарабанную полость (объем около 1 с м 3 см3 ), три слуховые косточки и слуховую (евстахиеву) трубу .

Рис. Среднее ухо
Барабанная полость находится в толщине височной кости, между барабанной перепонкой и костным лабиринтом. В барабанной полости помещаются слуховые косточки, мышцы, связки, сосуды и нервы. Стенки полости и все органы, находящиеся в ней, покрыты слизистой оболочкой.
В перегородке, отделяющей барабанную полость от внутреннего уха, находится два окна:
- овальное окно : находится в верхней части перегородки, ведет в преддверие внутреннего уха; закрыто основанием стремечка;
- круглое окно: расположено в нижней части перегородки , ведет в начало улитки; закрыто вторичной барабанной перепонкой.
В барабанной полости находятся три слуховые косточки: молоточек, наковальня и стремя (= стремечко) . Слуховые косточки имеют небольшие размеры. Соединяясь между собой, они образуют цепь, которая тянется от барабанной перепонки до овального отверстия. Все косточки соединяются между собой при помощи суставов и покрыты слизистой оболочкой.
Молоточек рукояткой сращен с барабанной перепонкой, а головкой при помощи сустава соединяется с наковальней , которая в свою очередь подвижно соединена со стременем . Основание стремени закрывает овальное окно преддверия.
Мышцы барабанной полости (натягивающая барабанную перепонку и стременная) удерживают слуховые косточки в состоянии напряжения и защищают внутреннее ухо от чрезмерных звуковых раздражений.
Слуховая (евстахиева) труба соединяет барабанную полость среднего уха с носоглоткой. Это мышечная трубка, которая раскрывается при глотании и зевании.
Слизистая оболочка, выстилающая слуховую трубу, является продолжением слизистой оболочки носоглотки, состоит из мерцательного эпителия с движением ресничек из барабанной полости в носоглотку.
Функции евстахиевой трубы:
- уравновешивание давления между барабанной полостью и внешней средой для поддержания нормальной работы звукопроводящего аппарата;
- защита от проникновения инфекций;
- удаление из барабанной полости случайно проникших частиц.
ВНУТРЕННЕЕ УХО
Внутреннее ухо состоит из костного и вставленного в него перепончатого лабиринта.
Костный лабиринт состоит из трех отделов: преддверия, улитки и трех полукружных каналов .
Преддверие - полость небольших размеров и неправильной формы, на наружной стенке которого расположены два окна (круглое и овальное), ведущие в барабанную полость. Передняя часть преддверия сообщается с улиткой через лестницу преддверия. Задняя часть содержит два вдавления для мешочков вестибулярного аппарата.
Улитка - костный спиральный канал в 2,5 оборота. Ось улитки лежит горизонтально и называется костным стержнем улитки. Вокруг стержня обвивается костная спиральная пластинка, которая частично перегораживает спиральный канал улитки и делит его на лестницу преддверия и барабанную лестницу . Между собой они сообщаются только через отверстие, находящееся у верхушки улитки.

Рис. Строение улитки: 1 - базальная мембрана; 2 - кортиев орган; 3 - рейснерова мембрана; 4 - лестница преддверия; 5 - спиральный ганглий; 6 - барабанная лестница; 7 - преддверно-завитковый нерв; 8 - веретено.
Полукружные каналы - костные образования, расположенные в трех взаимно перпендикулярных плоскостях. Каждый канал имеет расширенную ножку (ампулу).

Рис. Улитка и полукружные каналы
Перепончатый лабиринт заполнен эндолимфой и состоит из трех отделов:
- перепончатой улитки, или улиткового протока, продолжение спиральной пластинки между лестницей предверия и барабанной лестницей. В улитковом протоке находится слуховые рецепторы - спиральный, или кортиев, орган;
- трех полукружных каналов и двух мешочков , расположенных в преддверии, которые играют роль вестибулярного аппарата.
Между костным и перепончатым лабиринтом находится перилимфа --видоизмененная спинномозговая жидкость.
кортиев орган
На пластинке улиткового протока, которая является продолжением костной спиральной пластинки, находится кортиев (спиральный) орган .
Спиральный орган отвечает за восприятие звуковых раздражений. Он выполняет роль микрофона, трансформирующего механические колебания в электрические.
Кортиев орган состоит из опорных и чувствительных волосковых клеток.

Рис. Кортиев орган
Волосковые клетки имеют волоски, которые возвышаются над поверхностью и достигают покровной мембраны (мембраны тектория). Последняя отходит от края спиральной костной пластинки и свисает над кортиевым органом.
При звуковом раздражении внутреннего уха возникают колебание основной мембраны, на которой расположены волосковые клетки. Такие колебания вызывают розтяжение и сжатие волосков об покровную мембрану, и пораждают нервный импульс в чувствительных нейронах спирального ганглия.

Рис. Волосковые клетки
ПРОВОДНИКОВЫЙ ОТДЕЛ
Нервный импульс от волосковых клеток распространяется до спирального ганглия.
Затем по слуховому (преддверно-улитковому) нерву импульс поступает в продолговатый мозг.
В варолиевом мосту часть нервных волокон через перекрест (хиазму) переходит на противоположную сторону и идут в четверохолмие среднего мозга.
Нервные импульсы через ядра промежуточного мозга передаются в слуховую зону височной доли коры больших полушарий.
Первичные слуховые центры служат для восприятия слуховых ощущений, вторичные - для их обработки (понимание речи и звуков, восприятие музыки).

Рис. Слуховой анализатор
Лицевой нерв проходит вместе со слуховым нервом во внутреннее ухо и под слизистой оболочкой среднего уха следует к основанию черепа. Он может быть легко поврежден при воспалении среднего уха или травмах черепа, поэтому нарушения органов слуха и равновесия нередко сопровождаются параличом мимических мышц.
Физиология слуха
Слуховая функция уха обеспечивается двумя механизмами:
- звукопроведение : проведение звуков через наружное и среднее ухо к внутреннему уху;
- звуковосприятие : восприятие звуков рецепторами кортиева органа.
ЗВУКОПРОВЕДЕНИЕ
Наружное и среднее ухо и перилимфа внутреннего уха принадлежат к звукопроводящему аппарату, а внутреннее ухо, то есть спиральный орган и ведущие нервные пути – к звукоспринимающему аппарату. Ушная раковина благодаря своей форме концентрирует звуковую энергию и направляет ее в направлении к наружному слуховому проходу, который проводит звуковые колебания к барабанной перепонке.
Достигнув барабанной перепонки, звуковые волны вызывают ее колебание. Эти колебания барабанной перепонки передаются на молоточек, через сустав - на наковальню, через сустав - на стремя, которое закрывает окно преддверия (овальное окно). В зависимости от фазы звуковых колебаний, основа стремени то втискивается в лабиринт, то вытягивается из него. Эти движения стремени вызывают колебание перилимфы (см. рис.), которые передаются на основную мембрану улитки и на расположенный на ней кортиев орган.

В результате колебаний основной мембраны волосковые клетки спирального органа задевают нависающую над ними покровную (тенториальную) мембрану. При этом возникает растяжение или сжимание волосков, что и является основным механизмом превращения энергии механических колебаний в физиологичный процесс нервного возбуждения.
Нервный импульс передается окончаниями слухового нерва к ядрам продолгастого мозга. Отсюда импульсы проходят соответствующими ведущими путями к слуховым центрам в височных частях коры головного мозга. Здесь нервное возбуждение превращается в ощущение звука.

Рис. Путь звукового сигнала : ушная раковина - наружный слуховой проход - барабанная перепонка - молоточек - наковальня - стемечко - овальное окно - преддверие внутреннего уха - лестница преддверия - базальная мембрана - волосковые клетки кортиева органа. Путь нервного импульса : волосковые клетки кортиева органа - спиральный ганглий - слуховой нерв - продолговатый мозг - ядра промежуточного мозга - височная доля коры больших полушарий.
ЗВУКОВОСПРИЯТИЕ
Человек воспринимает звуки внешней среды с частотой колебаний от 16 до 20000 Гц (1 Гц = 1 колебание за 1 с).
Высокочастотные звуки воспринимаются нижней частью завитка, а низкочастотные звуки - его верхушкой.

Рис. Схематическое изображение основной мембраны улитки (указаны частоты, различимые разными участками мембраны)
Ототопика - с пособность определять местонахождение источника звука в случаях, когда мы не видим его, называется. Она связанная с симметричной функцией обоих ушей и регулируется деятельностью центральной нервной системы. Такая способность возникает потому, что звук, который идет сбоку, попадает в разные уши не одновременно: в ухо противоположной стороны - с опозданием в 0,0006 с, с другой интенсивностью и в другой фазе. Эти отличия восприятия звука разными ушами дают возможность определять направление источника звука.